Under heterosis understand the superiority of the first generation offspring over the parent forms (the best parent form) in terms of vitality, constitutional strength, endurance and productivity, which arises when crossing different races, breeds of animals and their zonal types ( E.K. Merkuryeva et al., 1991).
Some scientists believe heterosis effect and those cases when the offspring obtained as a result of crossing have indicators of economically useful traits that exceed the average between the parental forms, without exceeding the best of them.
Selection for heterosis is directly related to the theory and practice of breeding selection and selection and serves as one of the ways to increase animal productivity. Inbreeding is accompanied by inbreeding depression, increased homozygosity of inbred offspring and increased genetic similarity of the descendant to the ancestor. Heterosis has opposite biological and genetic properties.
The main indicators of heterosis are increasing embryonic and postembryonic viability; reduction of feed costs per unit of production; increasing early maturity, fertility, productivity; manifestation of greater opportunities for adaptation to changing conditions and new elements of technology. The wide range of the effect of heterosis, manifested in a variety of reactive characteristics, is a reflection of physiological and biochemical processes determined by the characteristics of the genetic apparatus of heterotic animals. One from its features is the greatest degree of expression only in the 1st generation of hybrids or crosses. Then heterosis imperceptibly fades and disappears in subsequent generations when hybrids are crossed with each other, unless special measures are taken to preserve the effect of heterosis.
The phenomenon of heterosis, or "hybrid vigor", was noticed in the practice of animal husbandry in ancient times, in particular in the production of mules by crossing a donkey with a mare. Hybrids from two-humped and one-humped camels (Batrian + dromedar = Nara and Maya), from a yak and a cow (khainaki) are also more powerful than the original forms. Darwin for the first time gave a scientific explanation of the “hybrid vigor” that occurs in offspring when unrelated organisms are crossed. He explained this effect by the biological dissimilarity of male and female gametes, which is caused by the influence of differences in the environment in which the parents live.
The term "heterosis" was introduced G. Schell (1914), who explained the presence of “hybrid vigor” by the state of heterozygosity in the genotype of an organism, formed as a result of crossing.
There have been many attempts to reveal the mechanism of the process of compatibility of hereditary factors that determines the effect of heterosis. Shell And East offered overdominance hypothesis. The heterosis hypothesis, formulated by Shell, East and Hayes, explains the phenomenon of heterosis by the presence of heterozygosity of various loci and the resulting overdominance, that is, when the effect of heterozygote Aa on the manifestation of the phenotype is stronger than the homozygous dominant genotype AA (that is, the effect of Aa is greater than the effect of AA) . That. its essence lies in the fact that high heterozygosity is better than homozygosity in providing diversity and strengthening of the physiological functions of the body. The significance of heterozygosity has been confirmed by works N.P. Dubinin, M. Lerner and other scientists.
Another explanation for heterosis, formulated Kiblom And Pellew (1910), is based on the fact that when crossing organisms carrying different homozygous genes in the genotype, for example AAbb and aaBB, in the crossbred offspring, recessive alleles pass into the heterozygous form of the AaBb genotype, in which the harmful effects of recessive genes are eliminated. The influence of dominant genes on the manifestation of heterosis can be explained by the simple cumulative effect of a large number of dominant genes, that is, there is an additive effect.
C. Davenport (1908) And D. Jones (1917) proposed to explain heterosis based on the hypothesis of the interaction of non-allelic dominant genes of both parents, which gives a total effect that causes heterosis. Kislovsky D.A. put forward to explain heterosis obligate heterozygosity hypothesis, according to which the body contains genes with dual effects - beneficial and harmful. In one direction this action is beneficial, in another it is neutral or even harmful to the body. In the process of evolution, those organisms survive in which the positive effect of genes was revealed in a heterozygous state, and the harmful effect turned out to be in a recessive state. Heterosis is primarily beneficial to the animal’s organism itself, which results from crossing. It follows that a high degree of heterozygosity - cause of heterosis.
To explain the phenomenon of heterosis, zootechnical theory has been put forward in recent years. concept of heterosis (A.I. Ovsyannikov, I.N. Nikitchenko and others) the essence of which is as follows:
- 1. The concept of contrasting crosses. According to it, the effectiveness of crossing is associated mainly with parental pairs that are contrasting and opposite in direction and body type. The principle of pair selection for heterosis is based on the proven effect of heterogeneous mating of queens with individual traits of productivity in combination with valuable traits of the father. A combination of rare extremes is not allowed (a comprehensive selection is in progress). Increasing vitality and hybrid vigor should be achieved by selecting breeds and individuals that differ in exterior, metabolism, and interior characteristics.
- 2. The principle of complementary action. The leading role in the formation of heterosis belongs to the combination of differences in the heredity of the original breeds. It has been determined that heterosis is a complex biological phenomenon in which 4 groups of factors are of decisive importance:
- a) direct effect of genes (level and number of productive qualities of the original breeds);
- b) maternal (reciprocal) effect;
- c) complementary action of genetic factors (additive), the action of dominant genes, the accumulation of which in the offspring during crossing enhances the development of the trait, causing heterosis;
- d) living conditions of the offspring of the first generation.
Various methods for producing heterotic animals have been tested in practice. These include: interspecific crosses, interbreed crosses, intrabreed crosses with heterogeneous selection, interline crosses, crosses of specially created inbred lines, mating of animals raised in different conditions. Each of these methods has its own characteristics and can be used to obtain heterosis not for all, but only for certain characteristics. Whatever methods are used to obtain heterosis, the individual characteristics of the manufacturer are of great importance. The more valuable its origin and the higher its ability to transmit its qualities to its offspring, the higher, other things being equal, will be the degree of manifestation of heterosis.
The huge role of heterosis in increasing productivity and improving other economically useful traits of animals prompted many scientists to find ways to consolidate it for a long time or at least preserve it for several generations. YES. Kislovsky was one of the first to theoretically substantiate the possibility of using heterosis in subsequent generations during interbreed variable crossing. He argued that with such crossing the features and positive aspects of absorption and industrial crossings are combined.
There are also other biochemical explanations for heterosis. It is believed that the main reason for hybrid power is the formation of sensitive copies of structural genes on chromosomes, which form an excess of information in cells and determine the high compatibility of metabolic processes ( Severin, 1967). Explanations for the heterosis effect can be found in the assumption that crossbreeds have polymorphic types of proteins (isoenzymes), which differ in some properties. The parental forms do not have enzyme polymorphism, and when they are crossed, polymorphism is formed in the crossbreeds and the number of polymorphic loci is therefore greater in them than in the parents. This, according to some scientists ( Fincham, 1968; Kirpichnikov, 1974), explains the overdominance effect.
F.M. Mukhametgaliev (1975) believes that the mutual stimulation of genomes during fertilization is equivalent to the additive effect of combined genetic systems and is the basis for the appearance of heterosis, but is not the cause of the emergence of new qualities in the genetic material, therefore heterosis manifests itself in quantitative changes in characteristics and has a polygenic type of inheritance.
A new approach to explaining the heterosis effect suggests V.G. Shakhbazov (1968). He believes that heterosis has a biophysical basis, since during fertilization an exchange of electrical charges of homologous chromosomes occurs, which increases the activity of chromosomes in hybrid zygotes. This leads to the accumulation of acidic proteins and RNA, increases the nucleolus-nuclear ratio and increases the rate of mitotic division.
The above explanations of the causes of the heterosis effect indicate a lack of unity in the scientific explanation of the phenomenon of heterosis, and therefore the problem remains for further study and consideration. Despite this, in animal husbandry practice, animal selection techniques are used to consolidate and enhance the effect of heterosis. There are several techniques for calculating the magnitude of the heterosis effect. There is a so-called true type of heterosis, which is determined by the magnitude of the superiority of the trait in crossbred animals over both parental forms. Another type of heterosis - hypothetical, when the characteristics of the crossbred offspring exceed the arithmetic mean level of the trait of both parents.
Professor N.A. Yurasov defines the line as a microbreed. From this point of view, a successful cross of lines can be considered as a result of the manifestation of heterosis. Just as heterosis during interbreeding is the result of the genetic diversity of the parents, heterosis during line crosses can be explained by the same phenomenon. The more carefully the crossed lines are selected and consolidated, the higher the effect can be expected.
Testing parental pairs and entire lines for compatibility to obtain heterosis is the basis of one of the effective techniques, which is called continuous reciprocal selection. Modern ideas about the causes of heterosis are based on the fact that heterosis is the result of the interaction of many genes. Their multiple actions lead to the heterosis effect. This explanation is called balance heterosis (Dobzhansky, 1952).
Revealed ecological type of heterosis (Merkuryeva, 1980), which is caused by the process of acclimatization and manifests itself in animals of the first ecological generation. This type of heterosis manifested itself in increased milk production of offspring born in the Ryazan region from Ayrshire cows imported from Finland. In subsequent generations, milk yield decreased to a level corresponding to the genetic potential of the introduced group of cows.
For the purpose of special use of heterosis in animal husbandry, methods of industrial and variable crossing have been developed. In such cases, purebred animals of two breeds are crossed, and their offspring, superior to the representatives of the original breeds, are used in whole or in large part for fattening and slaughtering for meat. Most examples of the manifestation of heterosis in animals of different species were noted during interbreeding. In beef cattle breeding, when crossing different breeds, crossbreeds of the first generation exceed the original breeds in live weight. In dairy cattle, the effect of heterosis is observed more often in the total amount of milk fat per lactation, especially when crossing cows of different breeds with Jersey bulls.
In the breeding of animals and plants, a special place is occupied by the phenomenon of hybrid power, or heterosis, which is as follows.
When crossing different races, animal breeds and plant varieties, as well as inbred lines, F 1 hybrids are often superior to the original parent organisms in a number of traits and properties. Crossing hybrids with each other leads to the attenuation of this effect in subsequent generations. Currently, heterosis has been established for all studied species.
Although the effect of heterosis has been known since ancient times, its nature still remains poorly understood. The first interpretation of the biological significance of heterosis in the evolution of animals and plants and an attempt to explain the mechanism of this phenomenon belonged to Charles Darwin. He systematized a large number of facts and himself conducted experiments to study heterosis. According to Charles Darwin, heterosis is one of the reasons for the biological usefulness of crossing in the evolution of species. Cross-fertilization is supported by natural selection precisely because it serves as a mechanism for maintaining the greatest heterosis,
A deep scientific analysis of the phenomenon of heterosis became possible only from the beginning of the 20th century. after the discovery of basic genetic patterns. When considering heterosis, we will touch upon its occurrence when crossing predominantly inbred lines of the same species, since in this case it is easier to elucidate its genetic mechanism.
Since the beginning of this century, systematic studies of crosses between inbred lines have been carried out on maize. At the same time, G. Schell showed that crossing some lines produces hybrid plants that are more productive in terms of grain and vegetative mass than the original lines and varieties. Experimental data are presented showing the low yield of inbred lines, a significant increase in yield in F 1 and a decrease in F 2 during self-pollination of F 1 plants.
Over the past 30 years, in a number of countries, including ours, sowing with hybrid seeds has become the main method of producing corn both for grain and for silage. To obtain hybrid seeds, first inbred lines are created from the best varieties that meet the requirements of a given climatic region (an inbred line is created within 5-6 years by self-pollination). When selecting lines, their qualities and properties that need to be obtained from the future hybrid organism are assessed. Inbreeding within lines cannot be effective unless it is accompanied by selection.
Having created a large number of inbred lines, they begin crossing between them. Interline hybrids of the first generation are assessed by heterosis effect; Based on this indicator, lines with the best combinational value are selected and then propagated on a large scale to produce hybrid seeds. Work on creating inbred lines and assessing their combinational value is constantly carried out in breeding institutions. The more valuable lines are created, the sooner the best hybrid combinations with the required combination of properties can be found.
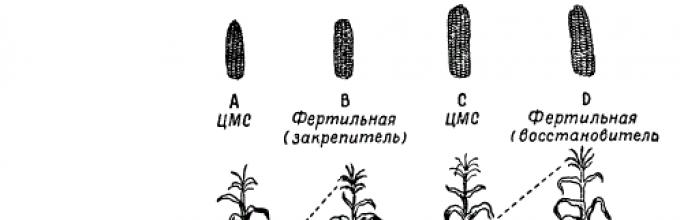
When obtaining hybrid seeds for production purposes, the original lines that give the greatest heterosis effect when crossed are sown in rows, alternating maternal and paternal forms. To ensure pollination between them, male inflorescences (panicles) are removed from the mother plants. Now a new scheme for the production of hybrid seeds using cytoplasmic male sterility has been developed, which has significantly reduced the labor required to remove panicles from maternal plants. In this way, simple interline corn hybrids are obtained. This method is, in principle, general for seed production of hybrids of various cross-pollinating plants.
At present, simple interline corn hybrids are not used in agricultural practice, since the costs of obtaining such seeds are not recouped. Sowing seeds of double interline hybrids is now being widely introduced into practice. The latter are obtained by crossing two simple hybrids exhibiting heterosis.

According to M.I. Khadzhinov and G.S. Galeev, the results of assessing the productivity of different hybrids - intervarietal, varietal and double interlinear - show that double interlinear hybrids are the most productive.

The selection of simple hybrids to obtain the most productive double hybrids is an important stage in their selection. The best results are obtained by crossing lines originating from different varieties. So, for example, if one simple hybrid is obtained from crossing inbred lines of two varieties A X B, and another from crossing lines of other varieties C x D, then a double hybrid (A X B) x (C X D) gives heterosis more often than if a double hybrid would be obtained from crossing simple hybrids descended from lines of the same variety: (A x A 1 x (A 2 x A 3) or (B x B 1) x (B 2 x B 3).
For the successful development of hybrid seed production, it is necessary to study, firstly, how long inbreeding should be carried out in order to obtain homozygous lines, and, secondly, to develop methods for more quickly assessing their combinational value.
All that has been said in relation to corn seems to be general for the production of hybrids in other higher cross-pollinating plants, as well as in animals. Currently, in poultry and pig farming in a number of countries, the crossing of inbred lines originating from the same or different breeds is widely used. It should be especially emphasized that the widespread use of hybrids in livestock breeding is possible only with a high level of breeding work and the presence of valuable breeds, it is clear that in most cases, inbred lines, on average, will always have lower performance than varieties. The presence of heterosis should be discussed only in the case when the interline hybrid is superior not only to the parents (lines), but also to the varieties or breeds from which these lines originated.
As we know, genes determine the properties of an organism at all stages of ontogenesis, starting from the moment of fertilization. The genes of the oocyte nucleus are capable of determining the properties of the cytoplasm of the egg even before fertilization. The nature of the implementation of the genotype changes depending on the properties of the cytoplasm of the zygote. Thus, the manifestation of heterosis in a hybrid also depends on the properties of the cytoplasm. The role of the cytoplasm in determining heterosis is proven as follows. When reciprocally crossing two lines A x B and B x A, heterosis for the same properties often appears in hybrids of only one of the crosses and does not appear in hybrids of the other.
The manifestation of heterosis is due to the individual development of the hybrid. During ontogenesis it is realized unevenly. At some stages of ontogenesis, heterosis manifests itself according to some characteristics, and at others - according to others or only according to some. Thus, at an early age, the same hybrid may exhibit heterosis in relation to the growth rate of individual parts of the body and increased resistance to diseases, but it may not exist, for example, in relation to resistance to unfavorable temperatures. Heterosis due to this property may appear later.
The manifestation of heterosis is also strongly influenced by environmental factors in which the hybrid organism develops. There is reason to believe that heterosis in terms of viability and resistance to diseases is better detected under environmental conditions unfavorable for development. Even Charles Darwin suggested that heterosis in hybrids is due to a wider norm of adaptive reactions. Research in recent years has shown that F 1 hybrids, in comparison with inbred lines, are characterized by a greater variety of metabolic processes, the participation of a larger number of different metabolites, growth substances and enzymes. Heterosis manifests itself not only in the whole organism, but also at the cellular level.
Elucidation of the genetic mechanisms of heterosis remains an unresolved problem. Currently, there are three hypotheses trying to explain the occurrence of heterosis:
- heterozygous state for many genes,
- interaction of dominant favorable genes,
- overdominance - a heterozygote is superior to homozygotes.
As we have already said, when crossing homozygous inbred lines, first-generation hybrids acquire a heterozygous state for many genes. In this case, the effect of harmful recessive mutant alleles is suppressed by the dominant alleles of both parents. Schematically, this can be represented as follows: one inbred line in a homozygous state has a recessive allele of one gene aaBB, and the second - another gene AAbb. Each of these recessive gene alleles in the homozygous state determines some deficiency that reduces the viability of the inbred line. When crossing lines aaBB X AAbb, the hybrid combines the dominant alleles of both genes (AaBb). F 1 hybrids will show not only heterosis, but also uniformity in the indicated genes. In F 2, the number of individuals with two dominant genes in the heterozygous state will be only 4/16, so not all individuals are heterotic. In further generations, the number of heterozygotes decreases, and the number of homozygotes increases. For these reasons, heterosis fades in subsequent generations. This is the scheme of one of the genetic hypotheses of heterosis.
We gave an example with two genes, but physiological properties are determined by a huge number of genes. In addition, it is incorrect to assume that dominant alleles always have a positive effect, and recessive ones have a negative effect on the viability of the organism and its productivity. Dominant wild-type alleles are more likely to have a beneficial effect than recessive ones. This can be explained by the fact that unfavorable dominant mutations are subject to more stringent selection - they are already eliminated in the zygote and at the embryonic stages, and only favorable ones are retained by selection. And since the very property of gene dominance evolves under the control of selection, wild-type alleles turn out to be more favorable for the organism. Therefore, the selection of dominant alleles in a hybrid combination can more likely ensure heterosis. If heterosis were caused by a simple set of dominant alleles present in the population, then this set would be easy to compose through a series of crosses and obtain heterotic combinations. It is very possible that some breeds and varieties were developed precisely by crossing and combining lines with a set of favorable dominant alleles. But so far it has not been possible to fix heterosis in the hybrid combination of F 1, that is, to obtain forms that do not split into F 2.
D. Jones proposed an addition to the stated hypothesis back in 1917. According to D. Jones, different genes that give heterosis in combination are located within the same group of linkages: for example, AbcdE - in one, and, accordingly, aBCDe - in another homologous chromosome. It is this circumstance, from the point of view of D. Jones, that makes it difficult to select completely homozygous forms in F 2 for favorable dominant genes that give the combination heterosis. To implement such a combination of dominant alleles, it is necessary that in this chromosome pair AbcdE//aBCDe a minimum of double crossing over occurs, which would lead to the emergence of a chromosome with only dominant ABCDE alleles. The low probability of simultaneous occurrence of such an event in several chromosomes is, according to D. Jones, the reason that complicates the consolidation of heterosis. The F 1 hybrid contains all these dominant alleles, and therefore it exhibits heterosis.
This is the scheme for explaining heterosis according to the second hypothesis - the hypothesis of the interaction of dominant favorable genes, which is sometimes also called the hypothesis of a set of cumulative dominant alleles, and heterosis itself is called mutational. This hypothesis is generally based on the idea of a simple summation of the effect of dominant alleles with a complementary effect.
The third hypothesis is based on the fact that the heterozygous state of alleles is superior to the homozygous one (AA< Aa >aa). Here we can assume a favorable transposition of wild-type and mutant alleles, which somehow enhances the effect of the genes. This explanation of heterosis is called the overdominance hypothesis.
None of the three hypotheses can be considered the only correct one. Now it is premature to give preference to one or the other. They will probably all turn out to be correct, but for different cases. It is likely that each of the mechanisms envisioned by these hypotheses plays a role in determining hybrid power. Heterosis is a complex phenomenon both in terms of the mechanism of its occurrence and its manifestation in ontogenesis.
Obviously, the final conclusion about the genetic mechanisms of heterosis can be made only after the picture of the interaction of genes in the genetic system at the biochemical and molecular levels is revealed. As we already know, suppressing the manifestation of mutations with the help of suppressors allows us to correct disturbances in the action of the gene and its alleles. It is possible that the phenomenon of heterosis is the total result of the work not of the main genes that determine the development of the characteristics of the organism, but of a set of suppressive genes in the genotype. An important point in the study of heterosis is the study of plasma relations.
The main task of using heterosis in breeding is to consolidate it, that is, to preserve the effect of heterosis during the reproduction of the hybrid. The solution to this problem is conceived in several aspects: firstly, the consolidation of heterosis by transferring the hybrid organism from sexual reproduction to apomictic, which is apparently possible for some plants; secondly, the consolidation of heterosis by transferring a diploid hybrid exhibiting heterosis to a polyploid state. In this case, the heterozygous combination of genes will persist longer.
In vegetatively propagated plants, the maintenance of valuable hybrid combinations obtained sexually is carried out by vegetative propagation (cuttings, grafting, tubers, etc.).
There are a number of other ways to maintain heterosis over a number of generations, but all of them have not yet been sufficiently developed and tested.
If you find an error, please highlight a piece of text and click Ctrl+Enter.
1.1. Under term heterosis in the broad sense of the word, all positive effects leading to the superiority of first generation hybrids (F 1) over parental forms are understood.
Heterosis(from the Greek heteroiosis - change, transformation), “hybrid strength”, acceleration of growth to increase in size, increase in vitality and fertility of first-generation hybrids compared to parental forms in various crossings of both animals and plants. Heterosis is fully manifested in the first hybrid generation. In the second and subsequent generations, G. usually fades out. In vegetatively propagated plants, for example, potatoes, sugar cane, etc., heterosis is persistently transmitted to the vegetative offspring, since all plants of the clone genotypically correspond to the original maternal individual.
Advantages of heterotic hybrids: high quality seeds, increased yield, resistance to many diseases, very early ripening.
Disadvantages of heterotic hybrids: high price of seeds, high requirements for agricultural technology, high genetic homogeneity of hybrids leads to a strong spread of diseases, impossibility of reproduction and collection of seeds.
The selection of heterotic hybrids has a large importance for agricultural production. These hybrids often exceed the yield of conventional open-pollinated varieties by 30% or more. In some cases, the heterosis effect reaches 50%. The phenomenon of heterosis is widely used in the breeding of corn, sorghum, sunflower, tomato, pumpkin, cucumber, watermelon, onion, cabbage, sugar beet, and ornamental plants. It has also begun to be used in the breeding of rice, wheat, and cotton.
1.2. FACTORS CONDITIONING HETEROSIS
For a long time, attempts were made to explain the phenomenon of heterosis by individual genetic factors. Several hypotheses have emerged.
So, overdominance hypothesis explains the manifestation of heterosis by the heterozygous state of hybrids.
The second classical hypothesis about the manifestation of heterosis, the so-called dominance hypothesis, proceeds from the fact that it is not the heterozygous state in itself, but the accumulation of dominant productivity alleles caused by crossing that leads to heterosis.
Many studies conducted after the appearance of these hypotheses have shown that The causes of heterosis are complex. Obviously, heterosis is due to the physiological balance of metabolic processes, which is achieved more easily by heterotic hybrids than by homozygous forms.
Based on this, it was put forward genetic balance concept. It turned out that nuclear-plasmic interactions can also affect the balance of hybrids determined by chromosomal genes, especially if the hybrid idiotype contains genetically different mitochondria, which can lead to increased respiration and enzymatic activity. Thus, heterosis is determined not only by the genotypic, but also by the plasmatic constitution of the hybrids.
1.3. TYPES OF HYBRIDS USED IN PRODUCTION
The following types of hybrids for industrial use are distinguished:
1) interline:
Simple - from crossing two self-pollinated lines;
Trilinear - from pollination of a simple interline hybrid with pollen of a self-pollinated line;
Double - from crossing two simple interline hybrids;
Complex interline hybrids - obtained with the participation of more than four self-pollinated lines;
2) variety linear:
Simple - from pollination of varieties with pollen of the line;
Complex - from pollination of the variety with pollen of a simple interline hybrid;
3) line-varietal- from pollination of a simple hybrid with pollen of the variety;
4) intervarietal hybrids- from crossing two varieties;
5) hybrid (synthetic) populations- obtained by mixing seeds of simple hybrids and other components” of their free cross-pollination.
The highest yield increases are achieved by hybrids obtained using self-pollinated lines.
1.4. GENERAL SCHEME FOR SELECTION OF HEGEROSIS HYBRIDS
Heterotic selection consists of several stages. Breeding work begins with the selection of source material from which self-pollinating lines are created. Then the combinative ability of these lines is studied, and those most suitable for combination are used as parent forms to create simple, double and other types of hybrids and to compile hybrid populations.
Heterosis is an increase in the viability of hybrids due to the inheritance of a certain set of alleles of various genes from their dissimilar parents. This phenomenon is the opposite of the results of inbreeding, or inbreeding, which leads to homozygosity. The increase in the viability of first-generation hybrids as a result of heterosis is associated with the transition of genes to a heterozygous state, while recessive lethal and semi-lethal alleles that reduce the viability of hybrids do not appear. Also, as a result of heterozygotation, several allelic variants of the enzyme can be formed, acting together more effectively than individually (in the homozygous state). The mechanism of action of heterosis has not yet been fully elucidated. The phenomenon of heterosis depends on the degree of relationship between the parental individuals: the more distant relatives the parental individuals are, the more pronounced the effect of heterosis is in first-generation hybrids. The phenomenon of heterosis was observed even before the discovery of Mendel’s laws by I. G. Köllreuter, the term “heterosis” (translated from Greek - change, transformation), in 1908 G. Shull described heterosis in corn. In plants (according to A. Gustafson), three forms of heterosis are distinguished: the so-called. reproductive heterosis, as a result of which the fertility of hybrids and productivity increases, somatic heterosis, which increases the linear dimensions of the hybrid plant and its weight, and adaptive heterosis (also called adaptive), which increases the adaptability of hybrids to the action of unfavorable environmental factors. There are five main forms of manifestation of heterosis based on economically useful traits used in animal husbandry;
1. Hybrids and crosses of the first generation exceed their parents in live weight and viability.
2. Crossbreeds of the first generation occupy an intermediate position in live weight, but are noticeably superior to their parents in fertility and viability.
3. First generation hybrids are superior to their parents in constitutional strength, longevity, physical performance, with complete or partial loss of fertility.
4. Each individual trait behaves according to an intermediate type of inheritance, and in relation to the final product, typical heterosis is observed.
5. Hybrids (crossbreeds) do not exceed the best parental form in production, but have a higher level than the arithmetic average of both parents.
The manifestation of heterosis during crossing and the results of the latter are influenced by a number of factors; original breeds and their compatibility, maternal and paternal heredity, feeding and maintenance conditions. The success of crossbreeding depends primarily on the correct choice of breeds and their combining ability. Not all breeds can effectively cross with each other and produce the desired crossbred offspring. Only well-selected and compatibility-tested breeds are capable of passing on their valuable qualities to cross-breed descendants when crossed. Each breed differs from the other in its gene pool, i.e. a set of those genes that determine the level of productivity, external forms, physiological and anatomical characteristics of individuals of a given breed. Breed compatibility is the correspondence of the gene pool of one breed to the gene pool of another. Currently, in animal husbandry there are no sufficiently reliable methods that would allow us to predict in advance the most successful combinations.
All that remains is to test the compatibility of different breeds and transfer the most successful ones to practice. It is also difficult to decide on the choice of maternal and paternal breeds, although during crossing significant differences have been established between crosses from direct and reverse mating. Many note the predominant influence of the mother’s body on the inheritance of economically useful traits in offspring. The maternal effect is determined by cytoplasmic heredity and the influence of the maternal organism as a nutrient medium on the embryo during the fruiting period. However, there are examples of the predominance of influence on certain characteristics of paternal heredity, which is difficult to explain. Although it can definitely be argued that for the success of crossing, individual selection of sires is no less important than in purebred breeding. The conditions of keeping animals have a great influence on the characteristics of the offspring. It is known that crossbreeds with combined heredity react much more strongly to environmental changes than purebred animals. Therefore, feeding and maintenance, along with genetic characteristics, determine the results of crossing. Thus, when low-productive aboriginal cattle were mated with cultivated breeds under poor feeding conditions, the traits of the former dominated, i.e. the effect of crossing was negative. At the same time, intensive feeding of crossbred young animals helps to increase live weight by 20–30% and correct a number of exterior defects. So, the success of crossing depends on a whole range of zootechnical measures, including a skillful selection of source breeds, selection of parent pairs, organization of adequate feeding of both the parents and the offspring obtained from them.
The practice of interbreed and interline hybridization indicates that it has not yet been possible to obtain guaranteed heterosis for specific paternal forms, which requires a large number of analyzing crosses. Therefore, modern breeding programs provide for the creation of a complex of specialized, combined lines, intra-breed types to obtain heterosis in hybrids in terms of productivity and reproductive qualities. The effectiveness of such selection is theoretically highest for traits with low heritability and with a large proportion of genes exhibiting dominance and overdominance. Despite the fact that sometimes the increase in productivity as a result of heterosis reaches 15%, the use of this phenomenon is accompanied by inadequate costs for the creation and preservation of original lines and breeds, overcoming inbreeding depression, conducting compatibility tests, and ensuring separate maintenance of individual breeds. It should be taken into account that selection to maintain compatibility must be carried out constantly and the original parental forms must be propagated for each generation of hybrids. The basis for the development of breeding programs designed to use heterosis over one generation is the reduction of the effect of heterosis during the subsequent breeding of hybrid individuals as a result of segregation of the original parental forms and loss of heterozygosity.
The idea of heterosis as the ability of individuals of the first generation to surpass the best of the parental forms does not contribute to the study of heterosis in the second generation. Obviously, the attenuation of heterosis during the breeding of crossbreeds is associated with a change in the frequency of the original genotypes. The genetically classical method of obtaining multiple heterosis is based on variable crosses. But it has disadvantages - it requires the reproduction of pure lines and, with an increase in the number of lines or breeds of more than three, it leads to complex rotational schemes, the implementation of which requires a long time. A significant drawback is that the same breeds act as maternal or paternal breeds, and this is undesirable in the case of specialized breeds. Considering that the problem of obtaining multiple heterosis is still being actively worked on, it makes sense to consider theoretical approaches to it. It is necessary to take into account that the main prerequisite for obtaining multiple heterosis is the need to reduce the cost of heterotic selection by fixing heterosis in several generations in the offspring of a certain heterozygosity. Multiple heterosis includes all cases when it is used beyond the first generation of hybrids and its effect persists for up to 3–4 generations. When studying the possibilities of obtaining such a phenomenon in animal husbandry, we will outline only theoretical ways of its implementation based on parthenogenesis, polyploidy, and genetic cloning. (Azimov A. 1997.)
When crossing certain varieties of a crop, F1 hybrids often differ from the parent forms in more powerful growth, better viability, higher productivity, resistance to diseases and sudden weather fluctuations. This property of first generation hybrids is called heterosis.
Heterosis is the property of first-generation hybrids to surpass their parents or the best of the parental forms in certain biological and economically valuable traits and properties, according to the degree of their expression.
The manifestation of heterosis depends on the type of gene pool of the population, the physiological state of the population at a certain stage of development, on climate conditions, season, solar insolation, nutrition, density of sowing or planting, intensity of the infectious background or insect pests or other factors and circumstances.
True (positive) heterosis manifests itself in a change in the severity of a trait in a hybrid towards an increase in comparison with the parent forms (for example, an increased content of vitamins, proteins, biologically active substances, improved shelf life of products, etc.). However, when crossing, depressive phenomena may occur in the offspring of a decrease in the indicator for some trait of the best parent. This change in the severity of the trait towards weakening occurs with negative heterosis.
Heterosis can cover different signs and properties of an organism. A. Gustafson proposed to distinguish between three main types of heterosis: somatic, reproductive and adaptive. Somatic heterosis - more powerful development of vegetative organs of hybrids; reproductive - more powerful development of reproductive organs, increased fertility, high yield of seeds and fruits; adaptive heterosis - increasing the adaptability of hybrids to changing environmental conditions and their competitiveness in the struggle for existence.
Heterotic hybrids of the first generation F1 exhibit increased viability, development power, especially strong growth ability, greater early maturity, productivity and uniformity, increased disease resistance, high marketability, improved fruit quality (increased dry matter content).
Heterosis can manifest itself in changes in physiological characteristics - increased cold resistance, drought resistance, improved shelf life, and general resistance to pathogens.
Heterosis is manifested in faster growth of plants in the initial and subsequent phases, in the accelerated formation of new leaves, in an increase in the size of each leaf and the total leaf surface area, in the power of the root system, etc. Heterosis is also manifested in such characteristics as the height of inflorescences, the number inflorescences on the main stem and side shoots, the number of flowers in the inflorescence, etc.
Heterosis is observed during crossings between varieties, as well as between species and forms that are genetically and ecologically distant. It manifests itself most strongly and can be controlled when crossing self-pollinated lines. Incineration makes it possible to decompose a variety-population into its constituent biotypes (lines).
The manifestation of heterosis is observed throughout the entire life cycle of the plant from the moment until the end of seed formation.
This or that type of manifestation of heterosis and the degree of variability in the expression of the trait mainly depend on the selection of crossed varieties. Typically, heterosis is more pronounced the more the crossed forms differ in morphological, biological, physiological and other characteristics. Heterosis is especially pronounced if the crossed varieties belong to different groups in terms of early ripening, method of cultivation, if they are zoned in different climatic zones or cultivated at different times of the year, when crossing inbred lines of cross-pollinating plants obtained as a result of self-pollination over several generations.
One of the reasons for the effect of heterosis is the elimination of the harmful effects of recessive genes in a heterozygous organism. Another mechanism of heterosis is the combination in a hybrid of favorable dominant genes present in the body of the parents and combined in the hybrid. Heterosis can also occur due to the fact that some genes manifest themselves more favorably when they are in a heterozygous state in the body.
THE IMPORTANCE OF SELECTION FOR HETEROSIS
Selection for heterosis refers to the creation of first-generation hybrids, characterized by high heterosis in yield, product quality and other economically important characteristics. Unlike combination breeding, in which crosses are carried out at the beginning of the breeding process to create genetic variability for selection, in the selection of heterotic hybrids, crossing serves for the mass production of seeds and their further practical use in production and represents the last stage of the breeding process. Selection of heterotic hybrids is of great importance for agricultural production. These hybrids often exceed the yield of conventional open-pollinated varieties by 30% or more. In some cases, the heterosis effect reaches 50%.