Мышечное сокращение является жизненно важной функцией организма, связанной с оборонительными, дыхательными, пищевыми, половыми, выделительными и другими физиологическими процессами. Все виды произвольных движений – ходьба, мимика, движения глазных яблок, глотание, дыхание и т. п. осуществляются за счет скелетных мышц. Непроизвольные движения (кроме сокращения сердца) – перистальтика желудка и кишечника, изменение тонуса кровеносных сосудов, поддержание тонуса мочевого пузыря – обусловлены сокращением гладких мышц. Работа сердца обеспечивается сокращением сердечной мускулатуры.
Структурная организация скелетной мышцы
Мышечное волокно и миофибрилла (рис. 1). Скелетная мышца состоит из множества мышечных волокон, имеющих точки прикрепления к костям и расположенных параллельно друг другу. Каждое мышечное волокно (миоцит) включает множество субъединиц – миофибрилл, которые построены из повторяющихся в продольном направлении блоков (саркомеров). Саркомер является функциональной единицей сократительного аппарата скелетной мышцы. Миофибриллы в мышечном волокне лежат таким образом, что расположение саркомеров в них совпадает. Это создает картину поперечной исчерченности.
Саркомер и филламенты. Саркомеры в миофибрилле отделены друг от друга Z -пластинками, которые содержат белок бета-актинин. В обоих направлениях от Z -пластинки отходят тонкие актиновые филламенты. В промежутках между ними располагаются более толстые миозиновые филламенты .
Актиновый филламент внешне напоминает две нитки бус, закрученные в двойную спираль, где каждая бусина – молекула белкаактина . В углублениях актиновых спиралей на равном расстоянии друг от друга лежат молекулы белка тропонина , соединенные с нитевидными молекулами белка тропомиозина.
Миозиновые филламенты образованы повторяющимися молекулами белка миозина . Каждая молекула миозина имеет головку ихвост . Головка миозина может связываться с молекулой актина, образуя так называемый поперечный мостик .
Клеточная мембрана мышечного волокна образует инвагинации (поперечные трубочки ), которые выполняют функцию проведения возбуждения к мембране саркоплазматического ретикулума. Саркоплазматичекий ретикулум (продольные трубочки) представляет собой внутриклеточную сеть замкнутых трубочек и выполняет функцию депонирования ионов Са++ .
Двигательная единица. Функциональной единицей скелетной мышцы является двигательная единица (ДЕ) . ДЕ – совокупность мышечных волокон, которые иннервируются отростками одного мотонейрона. Возбуждение и сокращение волокон, входящих в состав одной ДЕ, происходит одновременно (при возбуждении соответствующего мотонейрона). Отдельные ДЕ могут возбуждаться и сокращаться независимо друг от друга.
Молекулярные механизмы сокращения скелетной мышцы
Согласно теории скольжения нитей , мышечное сокращение происходит благодаря скользящему движению актиновых и миозиновых филламентов друг относительно друга. Механизм скольжения нитей включает несколько последовательных событий.
Головки миозина присоединяются к центрам связывания актинового филламента (рис. 2, А).
Взаимодействие миозина с актином приводит к конформационным перестройкам молекулы миозина. Головки приобретают АТФазную активность и поворачиваются на 120 ° . За счет поворота головок нити актина и миозина передвигаются на «один шаг» друг относительно друга (рис. 2, Б).
Рассоединение актина и миозина и восстановление конформации головки происходит в результате присоединения к головке миозина молекулы АТФ и ее гидролиза в присутствии Са++ (рис. 2, В).
Цикл «связывание – изменение конформации – рассоединение – восстановление конформации» происходит много раз, в результате чего актиновые и миозиновые филламенты смещаются друг относительно друга, Z -диски саркомеров сближаются и миофибрилла укорачивается (рис. 2, Г).
Сопряжение возбуждения и сокращения в скелетной мышце
В состоянии покоя скольжения нитей в миофибрилле не происходит, так как центры связывания на поверхности актина закрыты молекулами белка тропомиозина (рис. 3, А, Б). Возбуждение (деполяризация) миофибриллы и собственно мышечное сокращение связаны с процессом элетромеханического сопряжения, который включает ряд последовательных событий.
В результате срабатывания нейромышечного синапса на постсинаптической мембране возникает ВПСП, который генерирует развитие потенциала действия в области, окружающей постсинаптическую мембрану.
Возбуждение (потенциал действия) распространяется по мембране миофибриллы и за счет системы поперечных трубочек достигает саркоплазматического ретикулума. Деполяризации мембраны саркоплазматического ретикулума приводит к открытию в ней Са++ -каналов, через которые в саркоплазму выходят ионы Са++ (рис. 3, В).
Ионы Са++ связываются с белком тропонином. Тропонин изменяет свою конформацию и смещает молекулы белка тропомиозина, которые закрывали центры связывания актина (рис. 3, Г).
К открывшимся центрам связывания присоединяются головки миозина, и начинается процесс сокращения (рис. 3, Д).
Для развития указанных процессов требуется некоторый период времени (10–20 мс). Время от момента возбуждения мышечного волокна (мышцы) до начала ее сокращения называют латентным периодом сокращения .
Расслабление скелетной мышцы
Расслабление мышцы вызывается обратным переносом ионов Са++ посредством кальциевого насоса в каналы саркоплазматического ретикулума. По мере удаления Са++ из цитоплазмы открытых центров связывания становится все меньше и в конце концов актиновые и миозиновые филламенты полностью рассоединяются; наступает расслабление мышцы.
Контрактурой называют стойкое длительное сокращение мышцы, сохраняющееся после прекращения действия раздражителя. Кратковременная контрактура может развиваться после тетанического сокращения в результате накопления в саркоплазме большого количества Са++ ; длительная (иногда необратимая) контрактура может возникать в результате отравления ядами, нарушений метаболизма.
Фазы и режимы сокращения скелетной мышцы
Фазы мышечного сокращения
При раздражении скелетной мышцы одиночным импульсом электрического тока сверхпороговой силы возникает одиночное мышечное сокращение, в котором различают 3 фазы (рис. 4, А):
Латентный (скрытый) период сокращения (около 10 мс), во время которого развивается потенциал действия и протекают процессы электромеханического сопряжения; возбудимость мышцы во время одиночного сокращения изменяется в соответствии с фазами потенциала действия;
Фаза укорочения (около 50 мс);
Фаза расслабления (около 50 мс).
Режимы мышечного сокращения
В естественных условиях в организме одиночного мышечного сокращения не наблюдается, так как по двигательным нервам, иннервирующим мышцу, идут серии потенциалов действия. В зависимости от частоты приходящих к мышце нервных импульсов мышца может сокращаться в одном из трех режимов (рис. 4, Б).
Одиночные мышечные сокращения возникают при низкой частоте электрических импульсов. Если очередной импульс приходит в мышцу после завершения фазы расслабления, возникает серия последовательных одиночных сокращений.
При более высокой частоте импульсов очередной импульс может совпасть с фазой расслабления предыдущего цикла сокращения. Амплитуда сокращений будет суммироваться, возникнет зубчатый тетанус – длительное сокращение, прерываемое периодами неполного расслабления мышцы.
При дальнейшем увеличении частоты импульсов каждый следующий импульс будет действовать на мышцу во время фазы укорочения, в результате чего возникнет гладкий тетанус – длительное сокращение, не прерываемое периодами расслабления.
Оптимум и пессимум частоты
Амплитуда тетанического сокращения зависит от частоты импульсов, раздражающих мышцу. Оптимумом частоты называют такую частоту раздражающих импульсов, при которой каждый последующий импульс совпадает с фазой повышенной возбудимости (рис. 4, A) и соответственно вызывает тетанус наибольшей амплитуды. Пессимумом частоты называют более высокую частоту раздражения, при которой каждый последующий импульс тока попадает в фазу рефрактерности (рис. 4, A), в результате чего амплитуда тетануса значительно уменьшается.
Работа скелетной мышцы
Сила сокращения скелетной мышцы определяется 2 факторами:
Числом ДЕ, участвующих в сокращении;
Частотой сокращения мышечных волокон.
Работа скелетной мышцы совершается за счет согласованного изменения тонуса (напряжения) и длины мышцы во время сокращения.
Виды работы скелетной мышцы:
динамическая преодолевающая работа совершается, когда мышца, сокращаясь, перемещает тело или его части в пространстве;
статическая (удерживающая) работа выполняется, если благодаря сокращению мышцы части тела сохраняются в определенном положении;
динамическая уступающая работа совершается, если мышца функционирует, но при этом растягивается, так как совершаемого ею усилия недостаточно, чтобы переместить или удержать части тела.
Во время выполнения работы мышца может сокращаться:
изотонически – мышца укорачивается при постоянном напряжении (внешней нагрузке); изотоническое сокращение воспроизводится только в эксперименте;
изометричеки – напряжение мышцы возрастает, а ее длина не изменяется; мышца сокращается изометрически при совершении статической работы;
ауксотонически – напряжение мышцы изменяется по мере ее укорочения; ауксотоническое сокращение выполняется при динамической преодолевающей работе.
Правило средних нагрузок – мышца может совершить максимальную работу при средних нагрузках.
Утомление – физиологическое состояние мышцы, которое развивается после совершения длительной работы и проявляется снижением амплитуды сокращений, удлинением латентного периода сокращения и фазы расслабления. Причинами утомления являются: истощение запаса АТФ, накопление в мышце продуктов метаболизма. Утомляемость мышцы при ритмической работе меньше, чем утомляемость синапсов. Поэтому при совершении организмом мышечной работы утомление первоначально развивается на уровне синапсов ЦНС и нейро-мышечных синапсов.
Структурная организация и сокращение гладких мышц
Структурная организация. Гладкая мышца состоит из одиночных клеток веретенообразной формы (миоцитов ), которые располагаются в мышце более или менее хаотично. Сократительные филламенты расположены нерегулярно, вследствие чего отсутствует поперечная исчерченность мышцы.
Механизм сокращения аналогичен таковому в скелетной мышце, но скорость скольжения филламентов и скорость гидролиза АТФ в 100–1000 раз ниже, чем в скелетной мускулатуре.
Механизм сопряжения возбуждения и сокращения. При возбуждении клетки Cа++ поступает в цитоплазму миоцита не только из саркоплазматичекого ретикулума, но и из межклеточного пространства. Ионы Cа++ при участии белка кальмодулина активируют фермент (киназу миозина), который переносит фосфатную группу с АТФ на миозин. Головки фосфорилированного миозина приобретают способность присоединяться к актиновым филламентам.
Сокращение и расслабление гладких мышц. Скорость удаления ионов Са++ из саркоплазмы значительно меньше, чем в скелетной мышце, вследствие чего расслабление происходит очень медленно. Гладкие мышцы совершают длительные тонические сокращения и медленные ритмические движения. Вследствие невысокой интенсивности гидролиза АТФ гладкие мышцы оптимально приспособлены для длительного сокращения, не приводящего к утомлению и большим энергозатратам.
Физиологические свойства мышц
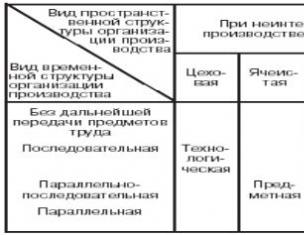
Общими физиологическими свойствами скелетных и гладких мышц являются возбудимость и сократимость . Сравнительная характеристика скелетных и гладких мышц приведена в табл. 6.1. Физиологические свойства и особенности сердечной мускулатуры рассматриваются в разделе «Физиологические механизмы гомеостаза».
Таблица 7.1. Сравнительная характеристика скелетных и гладких мышц
|
Свойство |
Скелетные мышцы |
Гладкие мышцы |
|
Скорость деполяризации |
медленная |
|
|
Период рефрактерности |
короткий |
длительный |
|
Характер сокращения |
быстрые фазические |
медленные тонические |
|
Энергозатраты | ||
|
Пластичность | ||
|
Автоматия | ||
|
Проводимость | ||
|
Иннервация |
мотонейронами соматической НС |
постганглионарными нейронами вегетативной НС |
|
Осуществляемые движения |
произвольные |
непроизвольные |
|
Чувствительность к химическим веществам | ||
|
Способность к делению и дифференцировке |
Пластичность гладких мышц проявляется в том, что они могут сохранять постоянный тонус как в укороченном, так и в растянутом состоянии.
Проводимость гладкой мышечной ткани проявляется в том, что возбуждение распространяется от одного миоцита к другому через специализированные электропроводящие контакты (нексусы).
Свойство автоматии гладкой мускулатуры проявляется в том, что она может сокращаться без участия нервной системы, за счет того, что некоторые миоциты способны самопроизвольно генерировать ритмически повторяющиеся потенциалы действия.
Введение
Вся жизнедеятельность животных и человека неразрывно связана с механическим движением, осуществляемым мышцами. Все телодвижения, кровообращение, дыхание и прочие акты возможны благодаря наличию в организме мышц, обладающих специальным белковым сократительным комплексом -- актомиозином.
Однако наличие сократительных элементов имеет значение не только при совершении вышеперечисленных макродвижений. В настоящее время накапливается все больше и больше данных о роли сократительных элементов в микропроцессах, в частности при активном транспорте веществ через мембраны и при движении цитоплазмы. Как было установлено, цитоплазма всех клеток находится в постоянном движении. По данным Камия, цитоплазма обладает колебательным, циркуляционным, фонтанирующим и другими видами движения, что, несомненно, играет большую роль в протекании метаболических процессов в клетках. В настоящее время нет единой точки зрения на причины происхождения этих движений цитоплазмы, однако наиболее вероятной является гипотеза функционирования сократительных элементов, подобных мышечным.
Сокращение скелетных мышц
мышца гладкая сокращение возбудимость
Основными физиологическими свойствами мышц являются их возбудимость, проводимость и сократимость. Последняя проявляется или в укорочении мышцы, или развитии напряжения.
Миография Для регистрации мышечного сокращения применяется методика миографии, т.е. графической регистрации сокращения с помощью рычажка, присоединенного к одному концу мышцы. Свободный конец рычажка чертит на ленте кимографа кривую сокращения - миограмму. Этот способ регистрации мышечного сокращения прост и не требует сложного оборудования, но имеет тот недостаток, что инерция рычажка и его трении по поверхности ленты кимографа несколько искажают запись. Во избежание этого недостатка теперь применяют специальный датчик, преобразующий механические изменения (линейные перемещения или усилия мышцы) в колебания силы электрического тока. Последние регистрируются с помощью шлейфного или катодного осциллографа.
Точной методикой является также оптическая регистрация, производимая с помощью пучка света, отраженного от зеркальца, наклеенного на брюшко мышцы.
По своим механическим свойствам мышцы относятся к эластомерам -- материалам, обладающим эластичностью (растяжимостью и упругостью). Если мышцу подвергнуть действию внешней механической силы, то она растягивается. Величина растяжения мышцы в соответствии с законом Гука будет пропорциональна величине деформирующей силы (в определенных пределах):
где Дl -- абсолютное удлинение мышцы; l -- начальная длина мышцы; F -- деформирующая сила; S -- площадь поперечного сечения мышцы; б -- коэффициент упругости. Величина отношения F/S называется механическим напряжением, а величина l/б -- модулем упругости; он показывает величину напряжения, необходимого для удлинения тела в 2 раза относительно начальной длины.
По своим свойствам мышца приближается к каучуку, модуль упругости для обоих этих материалов равен примерно 10 кгс/см 2 . Мышцы обладают и другими свойствами, присущими каучуку. Как и при растяжении каучука, при сильном растяжении мышцы наблюдается локальная кристаллизация (упорядочение макромолекулярной белковой структуры фибриллярного типа). Это явление было изучено методом рентгеноструктурного анализа. При этом освобождается кристаллизационное тепло, в результате чего температура мышцы при растяжении повышается.
После того как внешнюю силу убирают, мышца восстанавливает свою длину. Однако восстановление не бывает полным. Наличие остаточной деформации характеризует пластичность мышцы -- способность сохранять форму после прекращения действия силы. Таким образом, мышца не является абсолютно упругим телом, а обладает вязкоупругими свойствами. При очень сильном растяжении мышца ведет себя как нормальное упругое тело. В этом случае при растяжении температура мышцы понижается.
При сокращении мышцы развивается напряжение и совершается работа. Мышцы обладают сократительными и эластическими элементами. Поэтому возникающее напряжение и совершаемая работа обусловлены не только активным сокращением сократительного комплекса, но и пассивным сокращением, определяемым эластичностью или так называемым последовательным упругим компонентом мышцы. За счет последовательного упругого компонента работа совершается только в том случае, если мышца была предварительно растянута, и величина этой работы пропорциональна величине растяжения мышцы. Этим в большой степени объясняется то, что наиболее мощные движения совершаются при большой амплитуде, обеспечивающей предварительное растяжение мышц.
Мышечные сокращения делятся на изометрические - происходящие при неизменной длине мышцы, и изотонические - происходящие при неизменном напряжении. Чисто изометрические или чисто изотонические сокращения с большим или меньшим приближением можно получить только в лабораторных условиях при работе на изолированных мышцах. В организме сокращения мышц никогда не бывают чисто изометрическими или чисто изотоническими.
Скелетные мышцы с помощью сухожилий прикрепляются к костям, которые образуют систему рычагов. В большинстве случаев мышцы прикрепляются к костям так, что при их сокращении наблюдается выигрыш в амплитуде движений и эквивалентный проигрыш в силе. Плечо рычага мышцы в большинстве случаев бывает меньше плеча рычага соответствующей кости. Согласно Аккерману, механический выигрыш в амплитуде движений большинства конечностей человека имеет величину от 2,5 до 20. Для двуглавой мышцы плеча он равен приблизительно 10. При движении костей соотношение плеч рычагов мышц и костей меняется, что приводит к изменению напряжения мышц. По этой причине изотонических сокращений в естественных условиях не наблюдается. По этой же причине в процессе сокращения меняются вышеприведенные величины механического выигрыша в амплитуде движений.
В зависимости от величины силы, которую преодолевает мышца, скорость сокращения (укорочения) мышцы бывает различной. Хилл на основе опытных данных, полученных при работе на изолированных мышцах, вывел так называемое основное уравнение сокращения мышцы. Согласно Хиллу, скорость сокращения мышцы v находится в гиперболической зависимости от величины нагрузки F:
(F + a) (v + b) = const,
где а и b -- константы, приблизительно равные? F и соответственно? v.

Рис.1. Зависимость скорости сокращения мышцы лягушки от величины нагрузки
Байером были сделаны интересные замечания к уравнению. Уравнение приводится к виду
F" v" = const,
если принять F" = F + a и v" = v + b. Произведение F х v" представляет собой общую мощность, развиваемую мышцей при сокращении. Так как Fv меньше F"v", т. е. внешняя мощность меньше общей мощности, то следует предположить, что мышца совершает не только внешнюю работу, но еще и некоторую внутреннюю работу, проявляющуюся в том, что нагрузка как бы увеличивается на величину а, а скорость сокращения на величину b. Эту внутреннюю работу можно интерпретировать как потерю энергии на внутримолекулярное трение в форме теплового рассеивания. Тогда с учетом высказанных замечаний можно отметить, что общая мощность мышцы в физиологических пределах является постоянной величиной, не зависящей от величины нагрузки и скорости сокращения.
С термодинамической точки зрения мышца представляет собой систему, которая преобразует химическую энергию (энергию АТФ) в механическую работу, т. е. мышца является хемо-механической машиной.
Как уже отмечалось, при сокращении мышцы происходит теплообразование. Хиллом с помощью термоэлектрических методов было установлено, что при каждом раздражении вначале выделяется постоянная по величине и не зависящая от нагрузки теплота активации Q, а затем теплота сокращения kДl, пропорциональная сокращению мышцы Дl и не зависящая от нагрузки (k -коэффициент пропорциональности). Если сокращение изотоническое, то мышца производит работу А, равную произведению нагрузки F на величину сокращения: А = FДl. Согласно первому закону термодинамики, изменение внутренней энергии ДU мышцы будет равняться сумме выделенного тепла и совершенной работы:
-ДU = Q + kДl + FДl = Q + Дl (F + k)
Тогда КПД мышечного сокращения будет равен:

Учитывая, что величины Q и k не зависят от F, из последнего уравнения следует, что в определенных пределах КПД мышечного сокращения будет увеличиваться при увеличении нагрузки.
Хилл на основе полученных им в опытах данных определил, что КПД мышечного сокращения примерно равен 40%. Если бы мышца работала как тепловая машина с КПД 40%, то при температуре среды 20 0 С температура мышцы должна была бы быть равной 215 0 С. Величина КПД 40% показывает эффективность превращения энергии АТФ в механическую энергию. Если учесть, что КПД окислительного фосфорилирования, в процессе которого синтезируется АТФ, имеет величину около 50%, то полная эффективность превращения энергии питательных веществ в механическую энергию будет равна приблизительно 20%.
Методы раздражения мышц. Для того чтобы вызвать сокращение мышцы, ее подвергают раздражению. Непосредственное раздражение самой мышцы (например, электрическим током) называется прямым раздражением; раздражение двигательного нерва, ведущее к сокращению иннервированной этим нервом мышцы, называется непрямым раздражением. Ввиду того, что возбудимость мышечной ткани меньше, чем нервной, приложение электродов раздражающего тока непосредственно к мышце еще не обеспечивает прямого раздражения: ток, распространяясь по мышечной ткани, действует в первую очередь на находящиеся в ней окончания двигательных нервов и возбуждает их, что ведет к сокращению мышцы. Чтобы получить сокращение мышцы под влиянием прямого раздражения, необходимо либо выключить в ней двигательные нервные окончания ядом кураре, либо прикладывать стимул через введенный внутрь мышечного волокна микроэлектрод.
Сокращение мышц — это сложный процесс, состоящий из целого ряда этапов. Главными составляющими здесь являются миозин, актин, тропонин, тропомиозин и актомиозин, а также ионы кальция и соединения, которые обеспечивают мышцы энергией. Рассмотрим виды и механизмы мышечного сокращения. Изучим, из каких этапов они состоят и что необходимо для цикличного процесса.
Мышцы
Мышцы объединяются в группы, у которых одинаковый механизм мышечных сокращений. По этому же признаку они и разделяются на 3 вида:
- поперечно-полосатые мышцы тела;
- поперечно-полосатые мышцы предсердий и сердечных желудочков;
- гладкие мышцы органов, сосудов и кожи.
Поперечно-полосатые мышцы входят в опорно-двигательный аппарат, являясь его частью, так как помимо них сюда входят сухожилия, связки, кости. Когда реализуется механизм мышечных сокращений, выполняются следующие задачи и функции:
- тело передвигается;
- части тела перемещаются друг относительно друга;
- тело поддерживается в пространстве;
- вырабатывается тепло;
- кора активируется посредством афферентации с рецептивных мышечных полей.
Из гладких мышц состоит:
- двигательный аппарат внутренних органов, в который входят легкие и пищеварительная трубка;
- лимфатическая и кровеносная системы;
- система мочеполовых органов.
Физиологические свойства

Как и у всех позвоночных животных, в человеческом организме выделяют три самых важных свойства волокон скелетных мышц:
- сократимость — сокращение и изменение напряжения при возбуждении;
- проводимость — движение потенциала по всему волокну;
- возбудимость — реагирование на раздражитель посредством изменения мембранного потенциала и ионной проницаемости.
Мышцы возбуждаются и начинают сокращаться от идущих от центров. Но в искусственных условиях используют тогда может раздражаться напрямую (прямое раздражение) или через нерв, иннервирующий мышцу (непрямое раздражение).
Виды сокращений
Механизм мышечных сокращений подразумевает преобразование химической энергии в механическую работу. Этот процесс можно измерить при эксперименте с лягушкой: ее икроножную мышцу нагружают небольшим весом, а затем раздражают легкими электроимпульсами. Сокращение, при котором мышца становится короче, называется изотоническим. При изометрическом сокращении укорачивания не происходит. Сухожилия не позволяют при развитии укорачиваться. Еще один ауксотонический механизм мышечных сокращений предполагает условия интенсивных нагрузок, когда мышца укорачивается минимальным образом, а сила развивается максимальная.
Структура и иннервация скелетных мышц
В поперечно-полосатые скелетные мышцы входит множество волокон, находящихся в соединительной ткани и крепящихся к сухожилиям. В одних мышцах волокна расположены параллельно длинной оси, а в других они имеют косой вид, прикрепляясь к центральному тяжу сухожильному и к перистому типу.

Главная особенность волокна заключается в саркоплазме массы тонких нитей — миофибрилл. В них входят светлые и темные участки, чередующиеся друг с другом, а у соседних поперечно-полосатые волокна находятся на одном уровне — на поперечном сечении. Благодаря этому получается поперечная полосатость по всему волокну мышц.
Саркомером является комплекс из темного и двух светлых дисков, и он отграничивается Z-образными линиями. Саркомеры — это сократительный аппарат мышцы. Получается, что сократительное мышечное волокно состоит из:
- сократительного аппарата (системы миофибрилл);
- трофического аппарата с митохондриями, комплексом Гольджи и слабой ;
- мембранного аппарата;
- опорного аппарата;
- нервного аппарата.
Мышечное волокно разделяется на 5 частей со своими структурами и функциями и является целостной частью ткани мышц.
Иннервация
Этот процесс у поперечно-полосатых мышечных волокон реализуется посредством нервных волокон, а именно аксонов мотонейронов спинного мозга и головного ствола. Один мотонейрон иннервирует несколько волокон мышц. Комплекс с мотонейроном и иннервируемыми мышечными волокнами называют нейромоторной (НМЕ), или (ДЕ). Среднее число волокон, которые иннервирует один мотонейрон, характеризует величину ДЕ мышцы, а обратную величину называют плотностью иннервации. Последняя является большой в тех мышцах, где движения небольшие и «тонкие» (глаза, пальцы, язык). Малое ее значение будет, напротив, в мышцах с «грубыми» движениями (например, туловище).
Иннервация может быть одиночной и множественной. В первом случае она реализуется компактными моторными окончаниями. Обычно это характерно для крупных мотонейронов. (называющиеся в этом случае физическими, или быстрыми) генерируют ПД (потенциалы действий), которые распространяются на них.
Множественная иннервация встречается, к примеру, во внешних глазных мышцах. Здесь не генерируется потенциал действия, так как в мембране нет электровозбудимых натриевых каналов. В них распространяется деполяризация по всему волокну из синаптических окончаний. Это необходимо для того, чтобы привести в действие механизм мышечного сокращения. Процесс здесь происходит не так быстро, как в первом случае. Поэтому его называют медленным.
Структура миофибрилл
Исследования мышечного волокна сегодня проводятся на основе рентгеноструктурного анализа, электронной микроскопии, а также гистохимическими методами.
Рассчитано, что в каждую миофибриллу, диаметр которой составляет 1 мкм, входит примерно 2500 протофибрилл, то есть удлиненных полимеризованных молекул белков (актина и миозина). Актиновые протофибриллы в два раза тоньше миозиновых. В покое эти мышцы находятся так, что актиновые нити кончиками проникают в промежутки между миозиновыми протофибриллами.
Узкая светлая полоса в диске А свободна от актиновых нитей. А мембрана Z скрепляет их.
На миозиновых нитях есть поперечные выступы длиной до 20 нм, в головках которых находится порядка 150 молекул миозина. Они отходят биополярно, и каждая головка соединяет миозиновую с актиновой нитью. Когда происходит усилие актиновых центров на нитях миозина, актиновая нить приближается к центру саркомера. В конце миозиновые нити доходят до линии Z. Тогда они занимают собой весь саркомер, а актиновые находятся между ними. При этом длина диска I сокращается, а в конце он исчезает полностью, вместе с чем линия Z становится толще.
Так, по теории скользящих нитей, объясняется сокращение длины волокна мышцы. Теория, получившая название «зубчатого колеса», была разработана Хаксли и Хансоном в середине двадцатого века.

Механизм мышечного сокращения волокна
Главным в теории является то, что не нити (миозиновые и актиновые) укорачиваются. Длина их остается неизменной и при растяжении мышц. Но пучки тонких нитей, проскальзывая, выходят между толстыми нитями, уменьшается степень их перекрытия, таким образом происходит сокращение.
Молекулярный механизм мышечного сокращения посредством скольжения актиновых нитей заключается в следующем. Миозиновые головки соединяют протофибриллу с актиновой. При их наклонах происходит скольжение, двигающее актиновую нить к центру саркомера. За счет биполярной организации миозиновых молекул на обеих сторонах нитей создаются условия для скольжения актиновых нитей в разные стороны.
При расслаблении мышц миозиновая головка отходит от актиновых нитей. Благодаря легкому скольжению расслабленные мышцы растяжению сопротивляются гораздо меньше. Поэтому они пассивно удлиняются.
Этапы сокращения
Механизм мышечного сокращения кратко можно подразделить на следующие этапы:
- Мышечное волокно стимулируется, когда потенциал действия поступает от мотонейронов из синапсов.
- Потенциал действия создается на мембране мышечного волокна, а затем распространяется к миофибриллам.
- Совершается электромеханическое сопряжение, представляющее собой преобразование электрического ПД в механическое скольжение. В этом обязательно участвуют ионы кальция.
Ионы кальция

Для лучшего понимания процесса активации волокна ионами кальция удобно рассмотреть структуру актиновой нити. Длина ее составляет порядка 1 мкм, толщина — от 5 до 7 нм. Это пара закрученных ниток, которые напоминают мономер актина. Примерно через каждые 40 нм здесь находятся сферические тропониновые молекулы, а между цепями — тропомиозиновые.
Когда ионы кальция отсутствуют, то есть миофибриллы расслабляются, длинные тропомиозиновые молекулы блокируют крепление актиновых цепей и мостиков миозина. Но при активизации ионов кальция тропомиозиновые молекулы опускаются глубже, и участки открываются.
Тогда миозиновые мостики прикрепляются к актиновым нитям, а АТФ расщепляется, и сила мышц развивается. Это становится возможным за счет воздействия кальция на тропонин. При этом молекула последнего деформируется, проталкивая тем самым тропомиозин.
Когда мышца расслаблена, в ней на 1 грамм сырого веса содержится больше 1 мкмоль кальция. Соли кальция изолированы и находятся в особых хранилищах. В противном случае мышцы бы все время сокращались.
Хранение кальция происходит следующим образом. На разных участках мембраны клетки мышцы внутри волокна имеются трубки, через которые происходит соединение со средой вне клеток. Это система поперечных трубочек. А перпендикулярно ей находится система продольных, на концах которых — пузырьки (терминальные цистерны), расположенные в непосредственной близости к мембранам поперечной системы. Вместе получается триада. Именно в пузырьках хранится кальций.
Так ПД распространяется внутрь клетки, и происходит электромеханическое сопряжение. Возбуждение проникает в волокно, переходит в продольную систему, высвобождает кальций. Таким образом осуществляется механизм сокращения мышечного волокна.

3 процесса с АТФ
При взаимодействии обеих нитей при наличии ионов кальция немалая роль отводится АТФ. Когда реализуется механизм мышечного сокращения скелетной мышцы, энергия АТФ применяется для:
- работы насоса натрия и калия, который поддерживает постоянную концентрацию ионов;
- этих веществ по разные стороны мембраны;
- скольжения нитей, укорачивающих миофибриллы;
- работы насоса кальция, действующего для расслабления.
АТФ находится в клеточной мембране, нитях миозина и мембранах ретикулума саркоплазматического. Фермент расщепляется и утилизируется миозином.
Потребление АТФ
Известно, что миозиновые головки взаимодействуют с актином и содержат элементы для расщепления АТФ. Последняя активизируется актином и миозином при наличии ионов магния. Поэтому расщепление фермента происходит при прикреплении миозиновой головки к актину. При этом чем больше поперечных мостиков, тем скорость расщепления будет выше.
Механизм АТФ
После завершения движения молекула АФТ обеспечивает энергией для разделения участвующих в реакции миозина и актина. Миозиновые головки разделяются, АТФ расщепляется до фосфата и АДФ. В конце подсоединяется новая АТФ-молекула, и цикл возобновляется. Таковым является механизм мышечного сокращения и расслабления на молекулярном уровне.
Активность поперечных мостиков будет продолжаться лишь до тех пор, пока происходит гидролиз АТФ. При блокировке фермента мостики не станут снова прикрепляться.
С наступлением смерти организма уровень АТФ в клетках падает, и мостики остаются устойчиво прикрепленными к актиновой нити. Так происходит стадия трупного окоченения.

Ресинтез АТФ
Ресинтез возможно реализовать двумя путями.
Посредством ферментативного переноса от креатинфосфата фосфатной группы на АДФ. Так как запасов в клетке креатинфосфата намного больше АТФ, ресинтез реализуется очень быстро. В то же время посредством окисления пировиноградной и молочной кислот ресинтез будет осуществляться медленно.
АТФ и КФ могут исчезнуть полностью, если ресинтез будет нарушен ядами. Тогда и кальциевый насос прекратит работу, вследствие чего мышца необратимо сократится (то есть настанет контрактура). Таким образом, нарушится механизм мышечного сокращения.
Физиология процесса
Подытоживая вышесказанное, отметим, что сокращение волокна мышцы состоит в укорочении миофибрилл в каждом из саркомеров. Нити миозина (толстые) и актина (тонкие) связаны концами в расслабленном состоянии. Но они начинают скользящие движения друг навстречу к другу, когда реализуется механизм мышечного сокращения. Физиология (кратко) объясняет процесс, когда под влиянием миозина выделяется необходимая энергия для преобразования АТФ в АДФ. При этом активность миозина будет реализована лишь при достаточном содержании ионов кальция, накапливающихся в саркоплазматической сети.
Процессы мышечной работы представляют собой многоуровневый комплекс физиологических и биохимических функций, жизненно важных для полноценной работы человеческого организма. Внешне подобные процессы можно наблюдать на примерах произвольных движений при ходьбе, беге, изменении мимики и т. д. Однако они охватывают гораздо больший спектр функций, в числе которых также значится работа дыхательного аппарата, органов пищеварения и выделительной системы. В каждом случае механизм мышечных сокращений подкрепляется работой миллионов клеток, в которой задействуются химические элементы и физические волокна.
Структурная организация мышцы
Мышцы формируются множеством волокон ткани, которые имеют узлы крепления к костям скелета. Они располагаются параллельно и в процессе мышечной работы взаимодействуют между собой. Именно волокна при поступлении импульсов обеспечивают механизм мышечного сокращения. Кратко структуру мышцы можно представить как систему, состоящую из молекул саркомер и миофибрилла. Важно понимать, что каждое мышечное волокно образуется множеством субъединиц миофибрилл, располагающихся продольно по отношению друг к другу. Теперь стоит отдельно рассмотреть саркомеры и филаменты. Поскольку они играют важную роль в двигательных процессах.
Саркомеры и филаменты
Саркомеры представляют собой сегменты волокон, которые отделяются так называемыми Z-пластинами, содержащими бета-актинин. От каждой пластины отходят актиновые филаменты, а промежутки заполняются толстыми миозиновыми аналогами. Актиновые элементы, в свою очередь, похожи на ниточки бус, закрученных в двойную спираль. В этой структуре каждая бусинка является молекулой актина, а в участках с углублениями в спирали находятся молекулы тропонина. Каждая из этих структурных единиц формирует механизм сокращения и расслабления мышечного волокна, связываясь друг с другом. Ключевую роль в возбуждении волокон играет клеточная мембрана. В ней заключены поперечные трубочки-инвагинации, которые активизируют функцию саркоплазматического ретикулума - это и будет возбуждающий эффект для мышечной ткани.
Двигательная единица

Теперь стоит отойти от углубленной структуры мышцы и рассмотреть двигательную единицу в общей конфигурации скелетной мышцы. Это будет совокупность мышечных волокон, иннервируемых отростками мотонейрона. Работа ткани мышцы независимо от характера действия будет обеспечиваться волокнами, включенными в состав одной двигательной единицы. То есть при возбуждении мотонейрона срабатывает механизм мышечных сокращений в рамках одного комплекса с иннервируемыми отростками. Такое разделение на мотонейроны позволяет целенаправленно задействовать конкретные мышцы, не возбуждая без надобности соседние двигательные единицы. По сути, вся мышечная группа одного организма делится на сегменты мотонейронов, которые могут объединяться в работе над сокращением или расслаблением, а могут действовать разнопланово или поочередно. Главное, что они независимы друг от друга и работают только с сигналами своей группы волокон.
Молекулярные механизмы мышечной работы
В соответствии с молекулярной концепцией о скольжении нитей, работа мышечной группы и, в частности, ее сокращение реализуется в ходе скользящего действия миозинов и актинов. Реализуется сложный механизм взаимодействия этих нитей, в котором можно выделить несколько процессов:
- Центральная часть миозиновой нити соединяются со связками актинов.
- Достигнутый контакт актина с миозином способствует конформационному перемещению молекул последнего. Головки вступают в фазу активности и разворачиваются. Таким образом осуществляются молекулярные механизмы мышечного сокращения на фоне перестройки нитей активных элементов по отношению друг к другу.
- Затем происходит взаимное расхождение миозинов и актинов с последующим восстановлением головной части последних.
Весь цикл выполняется несколько раз, в результате чего происходит смещение вышеупомянутых нитей, а Z-сегменты саркомеров сближаются и укорачиваются.

Физиологические свойства работы мышц
Среди основных физиологических свойств мышечной работы выделяют сократимость и возбудимость. Эти качества, в свою очередь, обуславливаются проводимостью волокон, пластичностью и свойством автоматии. Что касается проводимости, то она обеспечивает распространение процесса возбудимости между миоцитами по нексусам - это специальные электропроводящие контуры, отвечающие за проведение импульса сокращения мышцы. Однако после сокращения или расслабления тоже совершается работа волокон.
За их спокойное состояние в определенной форме отвечает пластичность, определяющая сохранение постоянного тонуса, в котором на текущий момент находится механизм мышечного сокращения. Физиология пластичности может проявляться как в виде сохранения укороченного состояния волокон, так и в их растянутом виде. Интересно и свойство автоматии. Она определяет способность мышц входить в рабочую фазу без подключения нервной системы. То есть миоциты самостоятельно вырабатывают ритмически повторяющиеся импульсы для тех или иных действий волокон.
Биохимические механизмы мышечной работы
В работе мышц участвует целая группа химических элементов, среди которых кальций и сократительные белки наподобие тропонина и тропомиозина. На базе этого энергетического обеспечения и выполняются рассмотренные выше физиологические процессы. Источником же этих элементов выступает аденозинтрифосфорная кислота (АТФ), а также ее гидролиз. При этом запас АТФ в мышце способен обеспечивать сокращение мышцы лишь в течение доли секунды. Несмотря на это, волокна могут отвечать на нервные импульсы в постоянном режиме.
Дело в том, что биохимические механизмы мышечного сокращения и расслабления с поддержкой АТФ связаны с процессом выработки резервного запаса макроэрга в виде креатинфосфата. Объем этого резерва в несколько раз превышает запас АТФ и в то же время способствует его генерации. Также помимо АТФ энергетическим источником для мышцы может выступать гликоген. К слову, на мышечные волокна приходится около 75% всего запаса данного вещества в организме.
Сопряжение возбудительных и сократительных процессов

В спокойном состоянии нити волокон не взаимодействуют друг с другом посредством скольжения, так как центры связок закрываются молекулами тропомиозина. Возбуждение может иметь место только после электромеханического сопряжения. Данный процесс также делится на несколько этапов:
- При активации нейромышечного синапса на мембране миофибриллы формируется так называемый постсинаптический потенциал, накапливающий энергию для действия.
- Возбуждающий импульс благодаря системе трубок расходится по мембране и активизирует ретикулум. Этот процесс в итоге способствует снятию барьеров с каналов мембраны, по которым выпускаются ионы, связывающиеся с тропонином.
- Белок тропонин, в свою очередь, открывает центры связок актина, после чего становится возможным механизм мышечных сокращений, но для его начала также потребуется соответствующий импульс.
- Использование открывшихся центров начнется в момент, когда к ним присоединятся головки миозина по описанной выше модели.
Полный цикл этих операций происходит в среднем за 15 мс. Период от начальной точки возбуждения волокон до полного сокращения называется латентным.
Процесс расслабления скелетной мышцы
При расслаблении мышц происходит обратный перенос ионов Са++ с подключением ретикулума и кальциевых каналов. В процессе выхода ионов из цитоплазмы количество центров связки сокращается, в результате чего происходит разъединение актиновых и миозиновых филаментов. Иными словами, механизмы мышечного сокращения и расслабления подключают те же функциональные элементы, но оперируют ими разными способами. После расслабления может наступать процесс контрактуры, при котором отмечается устойчивое сокращение мышечных волокон. Это состояние может сохраняться до момента, пока не наступит очередное действие раздражающего импульса. Бывает и контрактура краткого действия, предпосылками для которой становится тетаническое сокращение в условиях скопления ионов с большими объемами.

Фазы сокращения
Когда мускулатура приводится в действие раздражающим импульсом сверхпороговой силы, происходит одиночное сокращение, в котором можно выделить 3 фазы:
- Уже упомянутый выше период сокращения латентного типа, в процессе которого волокна накапливают энергию для совершения последующих действий. В это время проходят процессы электромеханического сопряжения и открываются центры связок. На данной стадии подготавливается механизм сокращения мышечного волокна, который активизируется после распространения соответствующего импульса.
- Фаза укорочения - длится 50 мс в среднем.
- Фаза расслабления - также длится примерно 50 мс.
Режимы мышечного сокращения
Работа при одиночном сокращении была рассмотрена как пример «чистой» механики мышечных волокон. Однако в естественных условиях такая работа не совершается, поскольку волокна находятся в постоянном отклике на сигналы двигательных нервов. Другое дело, что в зависимости от характера этого отклика может происходить работа в следующих режимах:
- Сокращения возникают при пониженной частоте импульсов. Если электрический импульс распространяется после завершения расслабления, то следует серия одиночных актов сокращения.
- Высокая частота импульсных сигналов может совпадать с расслабляющей фазой предшествующего цикла. В этом случае амплитуда, в которой работал механизм сокращения мышечной ткани, будет суммироваться, что обеспечит длительное сокращение с неполными актами расслабления.
- В условиях повышения частоты импульсов новые сигналы будут действовать в периоды укорочения, что спровоцирует длительное сокращение, которое не будет прерываться расслаблениями.
Оптимум и пессимум частоты

Амплитуды сокращений определяются частотой импульсов, которые раздражают мышечные волокна. В этой системе взаимодействия сигналов и откликов можно выделить оптимум и пессимум частоты. Первым обозначается частота, которая в момент действия будет накладываться на фазу повышенной возбудимости. В таком режиме может активизироваться механизм сокращения мышечного волокна с большой амплитудой. В свою очередь, пессимум определяет более высокую частоту, импульс которой приходится на фазу рефрактерности. Соответственно, в этом случае амплитуда уменьшается.
Виды работы скелетной мышцы
Мышечные волокна могут осуществлять работу динамически, статически и динамически-уступающе. Стандартная динамическая работа является преодолевающей - то есть мышца в момент сокращения перемещает объекты или его составные части в пространстве. Статическое действие мышцы в некотором роде избавлено от нагрузок, поскольку в этом случае не предусматривается изменение его состояния. Динамически-уступающий механизм мышечного сокращения скелетной мышцы срабатывает, когда волокна функционируют в условиях растяжения. Потребность в параллельном растяжении также может быть обусловлена тем, что работа волокон предполагает выполнение операций со сторонними телами.
В заключение

Процессы организации мышечного действия подключают самые разные функциональные элементы и системы. В работе задействуется сложный комплекс участников, каждый из которых выполняет свою задачу. Можно видеть, как в процессе активации механизма мышечных сокращений срабатывают и косвенные функциональные блоки. Например, это касается процессов генерации энергетического потенциала для совершения работы или системы блокировки центров связок, через которые происходит соединение миозинов и актинов.
Основная же нагрузка приходится непосредственно на волокна, которые выполняют те или иные действия по командам двигательных единиц. Причем характер выполнения определенной работы может быть разным. На него будут влиять параметры направляемого импульса, а также текущее состояние мышцы.
МЫШЕЧНОЕ СОКРАЩЕНИЕ - совокупность процессов, изменяющих механическое состояние мышцы; проявляется в укорочении мышцы и развитии ею механического напряжения.
Изучение физиологии мышечной системы, в частности механизмов М. с., началось еще в 18 в. Так, Д. Бернулли выдвинул принцип мышечной механики, заключающийся в том, что высота М. с. при прочих равных условиях пропорциональна длине мышечных волокон. В середине 19 в. Э. Вебер развил идею возникновения М. с. в результате напряжения мышц. Он считал, что мышца является упругим телом, в к-ром силы упругого сопротивления (напряжения) возрастают с нагрузкой. Т. о., была показана зависимость между длиной и напряжением, т. е. что изменение напряжения обусловливает изменение длины мышцы и наоборот. В 20 в. начались исследования тонкой структуры мышечных элементов и биохим, изменений, происходящих при М. с. Было установлено, что в основе сокращения лежит взаимодействие белков актина и миозина, а основной сократительный белок - миозин - обладает аденозинтрифосфатазной активностью. В 50-х гг. Хансон и Хаксли (J. Hanson, H. E. Huxley), сформулировав гипотезу скольжения сократительных нитей, заложили основы совр, понимания ультраструктуры мышц и механизма М. с.
Основой сократительной деятельности мышцы является одиночное М. с., возникающее в ответ на нервный импульс. Графически (рис.) одиночное М. с. имеет вид волны с восходящей и нисходящей фазами. Первая фаза называется сокращением, вторая - расслаблением. Расслабление более продолжительно во времени, чем сокращение. Общее время одиночного М. с. составляет доли секунды и зависит от функц, состояния мышцы. Продолжительность М. с. уменьшается при умеренной работе и возрастает при утомлении. Одиночное М. с. изолированного волокна в условиях постоянной температуры подчиняется закону «все или ничего » (см.). Между потенциалом действия мышцы и началом одиночного М. с. имеется кратковременный промежуток - скрытый период сокращения, во время к-рого потенциал действия распространяется по всей мышце. При этом из саркоплазматического ретикулума происходит выделение ионов кальция в пространство между сократительными протофибриллами (нитями), изменяется эластичность мышцы. В конце скрытого периода сокращения в самой мышце протекают механохимиче-ские реакции активации сократительных элементов, результатом к-рых является изотоническое или изометрическое М. с. Изотоническим называется такое М. с., при к-ром мышца свободно укорачивается; при изометрическом М. с. длина мышцы остается постоянной (оба ее конца закреплены) и меняется лишь напряжение. В организме в нормальных условиях в чистом виде изотонического и изометрического М. с. не наблюдается.
Если к мышце, к-рая не успела полностью расслабиться от предыдущего сокращения, поступает новый нервный импульс, то второе М. с. накладывается на первое (явление так наз. суперпозиции М. с., или механической суммации). При ритмическом возбуждении мышца приходит в состояние непрерывного М. с. (тетануса). При условии, когда интервал между импульсами небольшой, возникает слитное тетаническое М. с., или гладкий тетанус (см.). Напряжение во время тетануса выше, чем при одиночном М. с., упругие компоненты растягиваются полностью, и мышца достигает максимального напряжения; в процессе одиночного М. с. энергия тратится гл. обр. на растяжение эластических компонентов, расположенных в мышце последовательно,- нитей актина и миозина, Z-пластинки (Z-зоны), соединений концевых саркомеров с сухожилиями и сухожилий.
Поперечнополосатые мышцы имеют два важнейших механических свойства, определяющих характер М. с. Первое известно как взаимоотношение длина - сила (длина - напряжение) мышцы. Суть его заключается в том, что для каждой мышцы может быть найдена длина, при к-рой она развивает максимальную силу (напряжение).
Второе свойство мышц - это взаимозависимость силы и скорости М. с.: чем тяжелее груз, тем медленнее его подъем, и чем больше приложенная сила, тем меньше скорость укорочения мышцы. При очень большой нагрузке М. с. становится изометрическим; в этом случае скорость сокращения равна нулю. Без нагрузки скорость М.С. наибольшая. Между этими (экстремальными) значениями скорость укорочения изменяется в зависимости от нагрузки. В организме условия для осуществления той или иной формы мышечной деятельности не всегда оптимальны по параметрам «длина - напряжение» и «сила - скорость». Поэтому при осуществлении того или иного вида М. с. в результате деятельности ц. н. с. селективно отбирается количество быстрых или медленных ней-ромоторных единиц. Диапазон скоростей М. с. достаточно велик - от долей секунды (скелетные мышцы) до минут (гладкие мышцы). Он определяется многими факторами. Так, волокна поперечнополосатых мышц имеют короткие саркомеры, много миофибрилл, обильную сар-котубулярную систему, одно или два нервных окончания; гладкие мышцы характеризуются малым количеством и неупорядоченным расположением миофибрилл, слаборазвитой саркотубулярной системой, низкой активностью миозиновой АТФ-азы.
М. с. скелетных мышц может быть вызвано одним нервным импульсом. Для возникновения М. с. гладкой мышцы требуется ритмическая стимуляция. Скорость расслабления скелетных и гладких мышц значительно различается, т. к. зависит от количества упругих элементов в мышце, длины волокон, скорости поглощения ионов кальция и т. д.
Скорость связывания ионов кальция гладкой мышцей в десять и более раз меньше, чем в поперечнополосатой. В состоянии покоя основная часть ионов кальция в мышечном волокне депонируется в саркоплазматичес-ком ретикулума. Он представляет собой защитную систему внутриклеточных трубочек и цистерн, окружающих каждую миофибриллу. В механизмах М. с. особую роль играет та часть ретикулума, к-рая расположена в области Z-пластинки.
Основной сократительной единицей скелетной мышцы является сар-комер - участок мышечного волокна, расположенный между Z-пластинками. В саркомере упорядоченно расположены толстые (миозино-вые) и тонкие (актиновые) нити, имеется система продольных и поперечных канальцев (трубочек). Система канальцев состоит из впячи-ваний сарколеммы, называемых поперечными или Т-канальцами, сети продольных канальцев саркоплазматического ретикулума и пузырьков (концевых цистерн) между миофибрилл амп. Комплекс образований, включающий Т-канальцы и прилегающие две концевые цистерны, называется триадной системой мышечного волокна. Толстые и тонкие нити имеют зоны перекрытия и соединяются поперечными мостиками. Тонкие нити присоединены к Z-пластинкам.
При М. с. возбуждение мембраны мышечного волокна передается внутрь по канальцам Т-системы, стенки к-рых содержат заряженные частицы и проводят электрический сигнал (процесс перехода от возбуждения к сокращению называется электромеханической связью). Из саркоплазматического ретикулума под влиянием электрического сигнала, поступившего в T-систему, выходят ионы кальция и немедленно вступают во взаимодействие с тропо-нином, вследствие чего система тропонин - тропомиозин утрачивает способность тормозить взаимодействие актина с миозином (тропонин и тропомиозин - это белки-регуляторы, препятствующие взаимодействию актина и миозина). АТФ-аза миозина активируется актином при участии ионов магния. Энергия для М. с. освобождается активированной АТФ-азой актомиозина, гидролизующей АТФ на АДФ и фосфат. При этом освобождается большое количество энергии (до 10 ккал на 1 моль АТФ). Ресинтез АТФ из АДФ и фосфата происходит за счет креатинфосфата, процессов гликогено-лиза и гликолиза (см. Мышечная ткань, биохимия). Прекращение возбуждения ведет к снижению концентрации кальция в межфибрилляр-ном пространстве вследствие активного поглощения кальция мембранами саркоплазматического ретикулума. Энергия для активного поглощения ионов кальция высвобождается при расщеплении АТФ. Этот процесс по времени совпадает с теплопродукцией в фазу расслабления мышечного волокна. Связывание кальция мембранами ретикулума реактивирует систему тропонин - тропомиозин, к-рая вновь начинает тормозить взаимодействие актина и миозина (роль фактора расслабления, или фактора Марша,- вещества, обнаруженного в гомогенате свежей мышцы, понижающего АТФ-азную активность миозина и способствующего расслаблению, следует считать идентичной роли мембран саркоплазматического ретикулума).
Большинство теорий, объясняющих механизм М. с., основывается на идее Хаксли о взаимном скольжении толстых и тонких сократительных нитей. В скелетных мышцах максимальное напряжение развивается при полном взаимном перекрытии миози-новых и актиновых нитей в области образования мостиков. Когда мышца растянута до такой степени, что перекрытие отсутствует, напряжение падает до нуля. При сжатии мышцы тонкие нити, заходя друг за друга, нарушают процесс взаимодействия с толстьбми нитями, и напряжение мышцы падает. Скольжение нитей без изменения их длины, помимо образования поперечных мостиков и расщепления АТФ в молекуле миозина, возникает за счет каких-то конформационных изменений. По гипотезе Подольского (R. J. Podolsky) гидролиз АТФ меняет угол наклона мостиков, что приводит к вращению головок миозина или скручиванию миозиновой цепочки. В пользу этой гипотезы (т. е. гипотезы скольжения без изменения длины нитей) говорит тот факт, что напряжение в сокращенной мышце, если ее внезапно расслабить, восстанавливается в два этапа. Предполагают, что каждый мостик состоит из эластического плеча, к-рый, находясь в одном из нескольких энергетических стабильных положений, вступает во взаимодействие с акти-новой нитью. Напряжение восстанавливается в силу эластичности основания мостика и поворота миозиновой головки в положение, имеющее более низкий уровень потенциальной энергии. С физ.-хим. позиций М. с. можно представить как процесс перехода из жидкого состояния (комплекс молекул актина, миозина, АТФ, ионов кальция) в высокоэластическое (образование актомиозина, обладающего высокой эластичностью и упругостью).
Нарушения М. с. могут возникать при поражениях различных элементов нейромоторной единицы. Поражение тел мотонейронов или их аксонов, напр, при полиомиелите (см.) пли прогрессивной мышечной дистрофии (см. Миопатия), вызывает падение тонуса, атрофию или дистрофию мышц, мышечную фибрилляцию, нарушение чувствительности мышц к воздействию ацетилхолина, увеличение порогов к электрическому раздражению. Поражение моторной бляшки выражается слабостью и чрезвычайной утомляемостью мышц. Нарушение функции мышечной мембраны приводит к миотонии (см.), к-рая характеризуется нарушением нормального процесса расслабления: миотонические мышцы хорошо сокращаются, но не могут нормально расслабляться. Поражение функции собственно сократительного аппарата мышц наблюдается при прогрессирующей мышечной дистрофии и контрактурах. Нарушение сократительного аппарата наблюдается также и при мышечной гипертрофии, вызываемой чрезмерной нагрузкой какой-либо группы мышц.
Библиография: Арронет Н. И. Мышечные и клеточные сократительные (двигательные) модели, JI., 1971, библиогр.; Б ен-долл Дж. Мышцы, молекулы и движение, пер. с англ., М., 1970, библиогр.; Дещеревский В. И. Математические модели мышечного сокращения, М., 1977, библиогр.; Развитие сократительной функции мышц двигательного аппарата, под ред. JI. Г. Магазаника и Г, А. Наследо-ва, JI., 1974, библиогр.; Хилл А. В. Механика мышечного сокращения, пер. с англ., М., 1972; Close R.I. Dynamic properties of mammalian skeletal muscles, Physiol. Rev., v. 52, p. 129, 1972, bibliogr.; E b a s h i S. Excitation-contraction coupling, Ann. Rev. Physiol., v. 38, p. 293, 1976, bibliogr.